Salmonella Patojenite Adaları
Salmonella Pathogenicity Islands
Belgin SIRIKEN
Ondokuz Mayıs Üniversitesi Veteriner Fakültesi, Besin Hijyeni ve Teknolojisi Anabilim Dalı, Samsun.
Ondokuz Mayis University Faculty of Veterinary, Department of Food Hygiene and Technology, Samsun, Turkey.
ÖZET
Salmonella türleri fakültatif hücre içi yerleşim gösteren patojen bakterilerdir. Makrofajlara, dendritik ve epitelyal hücrelere invazyon yapabilmekte ve bütün Salmonella türleri patojen olarak kabul edilmektedir. İnvazyondan, hücre içinde canlılığını sürdürebilmesinden ve ekstraintestinal yayılımdan sorumlu olan virülans faktörlerini kodlayan genler Salmonella patojenite adaları (SPA) içinde yer alır. SPA'nın horizontal gen transferi ile kazanılmış olduğu düşünülmektedir. Bazı patojenite adaları Salmonella cinsi içinde korunmuş olarak bulunurken, bazıları belirli serotipler için özgüldür. SPA'nın varlığı ve özelliklerine göre Salmonella serotipleri arasında konak hücreye adaptasyon, virülans faktörleri ve oluşturdukları enfeksiyonların şiddeti açısından farklılıklar bulunmaktadır. Salmonella virülans gen kümeleri 12 patojenite adası içinde yerleşim göstermektedir. Enfeksiyonun intestinal fazını kapsayan virülans genleri SPA-1 ve SPA-2'de yer almakta; hücre içinde sağkalım, fimbriyal ekpresyon, çoklu antibiyotik direnci, magnezyum ve demir alımı ve sistemik enfeksiyon oluşumu için gerekli olan virülans faktörlerini kodlayan genler diğer SPA içinde bulunmaktadır. SPA dışında, alternatif sigma faktör ss (RpoS) regülatörü ile adaptif aside tolerans yanıtı (ATY) da Salmonella serotiplerinin diğer önemli virülans faktörleridir. Salmonella türlerinin virülan suşlarında bulunan RpoS ile ATY, enfeksiyonun intestinal fazı sırasında, mide asiditesi, safra tuzu, yetersiz oksijen, besin yetersizliği, antimikrobiyal peptidler, mukus ve doğal mikrobiyotanın varlığı gibi olumsuz çevresel koşullar altında canlılığını sürdürebilmesini ve aynı zamanda etkenin fagozom veya fagolizom gibi olumsuz koşullarda varlıklarını sürdürmesini sağlamaktadır. Bu derleme yazıda, Salmonella serotiplerinin virülans faktörlerini belirleyen önemli patojenite adaları ve virülans genlerinin regülasyonunda etkili bazı faktörler ile ilgili bilgiler özetlenmiştir.
Anahtar sözcükler: Salmonella; patojenite adası; sigma faktörü; adaptif aside tolerans yanıtı.
ABSTRACT
Salmonella species are facultative intracellular pathogenic bacteria. They can invade macrophages, dendritic and epithelial cells. The responsible virulence genes for invasion, survival, and extraintestinal spread are located in Salmonella pathogenicity islands (SPIs). SPIs are thought to be acquired by horizontal gene transfer. Some of the SPIs are conserved throughout the Salmonella genus, and some of them are specific for certain serovars. There are differences between Salmonella serotypes in terms of adaptation to host cell, virulence factors and the resulting infection according to SPA presence and characteristics. The most important Salmonella virulence gene clusters are located in 12 pathogenicity islands. Virulence genes that are involved in the intestinal phase of infection are located in SPI-1 and SPI-2 and the remaining SPIs are required for intracellular survival, fimbrial expression, magnesium and iron uptake, multiple antibiotic resistance and the development of systemic infections. In addition SPIs, Sigma ss (RpoS) factors and adaptive acid tolerance response (ATR) are the other two important virulence factors. RpoS and ATR found in virulent Salmonella strains help the bacteria to survive under inappropriate conditions such as gastric acidity, bile salts, inadequate oxygen concentration, lack of nutrients, antimicrobial peptides, mucus and natural microbiota and also to live in phagosomes or phagolysosomes. This review article summarizes the data related to pathogenicity islands in Salmonella serotypes and some factors which play role in the regulation of virulence genes.
Key words: Salmonella; pathogenicity island; sigma factor; adaptive acid tolerance response.
Geliş Tarihi (Received): 23.08.2012 • Kabul Ediliş Tarihi (Accepted): 06.09.2012
GİRİŞ
Enterobacteriaceae ailesi, Salmonella cinsinde yer alan bakteriler, Salmonella enterica (6 alt tür) ve Salmonella bongori (22 alt tür) olmak üzere iki tür içinde toplanmışlardır1. Salmonella türleri, tifo ve paratifo gibi enfeksiyonların yanı sıra bakteriyemi ve septisemi oluşturabilmekte; hafif ya da orta şiddette gıda zehirlenmelerine neden olmaktadırlar2. İnsanlarda izlenen salmonellozlarda etken olarak en sık S.enterica subsp enterica içinde yer alan S.Enteritidis, S.Typhimurium ve S.Newport serotipleri saptanmaktadır3.
Salmonella serotiplerinin konağa adaptasyonu, hücre içinde çoğalması, sağkalımı ve ekstraintestinal yayılımında çeşitli genler rol oynamakta ve bu genler aynı zamanda bakterinin virülans düzeyini de belirlemektedir4. Bilindiği gibi, patojen bakterilerin kromozomlarında "patojenite ile ilişkili adacıklar" adı verilen bölgede bulunan genler, bakterinin virülansını belirleyen çeşitli ürünleri kodlamaktadır5. Sadece patojen türlerde bulunan bu genomik bölgeler, horizontal gen transferi ile patojen olmayan türlere nakledildiğinde transfer olduğu bakteriye de kompleks virülans faktörleri kazandırmaktadır. Patojenite adaları çoğunlukla integrazlar, transpozonlar ve insersiyon sekans (IS) elementleri veya bu elementlerin bir kısmı gibi hareketli faktörleri kodlayan kriptik veya fonksiyonel genleri taşırlar. Salmonella genomu üzerinde bulunan patojenite adacıkları ise "Salmonella patojenite adaları" olarak adlandırılır4. Bu derleme yazıda, Salmonella serotiplerinin virülansında rol oynayan önemli patojenite adaları ile virülans genlerinin regülasyonunda etkin olan bazı faktörler özetlenmektedir.
SALMONELLA PATOJENİTE ADALARI
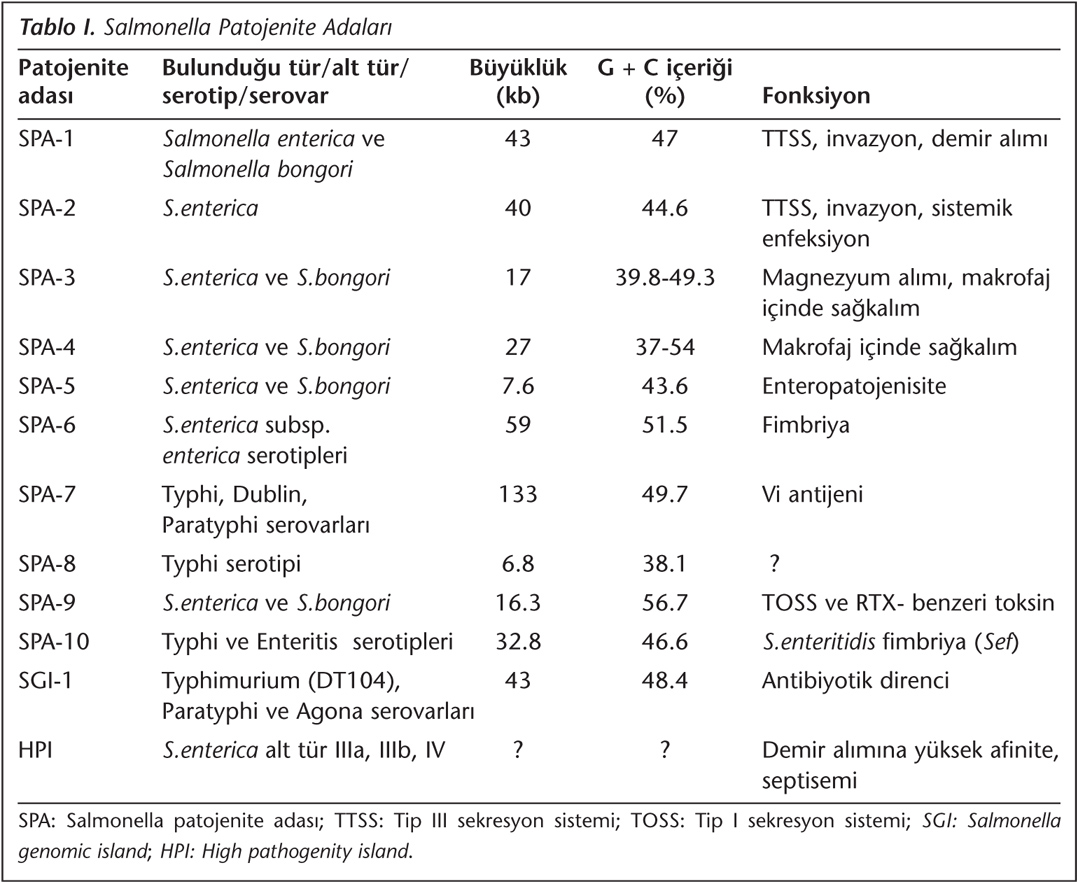
Salmonella patojenite adaları (SPA), çoğunlukla kromozom üzerinde bulunan 10-200 kilobaz (kb)'lık oldukça büyük genomik bölgeler olup, virülans faktörlerini kodlayan çok sayıda gen içermektedir4,6. Virülans faktörleri, bakterinin organizmaya adezyonu, kolonizasyonu, invazyonu, immün sistemden kaçışı ve toksin üretimi gibi özellikleri sağlayan ürünleri kapsamaktadır. Bu adacıklar sayesinde Salmonella türleri makrofajlar, dendritik ve epitelyal hücreleri enfekte edebilmektedir7,8,9. SPA'nın varlığı veya yokluğu Salmonella serotipleri arasında farklılıklar gösterir. Bazı patojenite adaları Salmonella cinsi içinde korunmuş olarak bulunurken, bazıları belirli serotipler için özgüldür; bu nedenle konakçı adaptasyonu, patojenite ve oluşturulan enfeksiyonun şiddeti açısından serotipler arasında farklılıklar bulunmaktadır4,6,7,10. Bugüne kadar birçok virülan Salmonella tipinde tanımlanan en önemli gen kümeleri 12 SPA içinde yer almaktadır4,11 (Tablo I).
Salmonella Patojenite Adası-1 (SPA-1)
SPA-1, 43 kb'lık segmente sahip olup, diğer patojenik bakterilerden horizontal gen transferi ile de geçebilmektedir. SPA-1, fagositoz yapmayan hücrelerin invazyonundan sorumlu 31 geni içerir; aynı zamanda kompleks Tip III sekresyon sistemi (TTSS) için gerekli proteinleri kodlar. Bu proteinler TTSS sistemi içinde bulunan invA, invB, invC, invF, invG, hilA, sipA, sipC, sipD, spar, orgA, sopB ve sopE genleri tarafından kodlanmaktadır. TTSS'nin dış membran proteini olan invG, protein sekresyonu ve bakterinin hücre içine alımında kritik rol oynar. InvA bir iç membran proteini olup, polipeptidlerin dışarı atılmasını sağlayan kanalların şekillenmesini sağlar. InvH ve HilD aksesuvar proteinler olup, bakterinin adezyonunda rol oynar. TTSS tarafından salgılanan iki protein alt grubu vardır. Bunlardan birinci grupta TTSS'ye polipeptid sekresyonunu sağlayan proteinler (InvJ ve SpaO) yer alırken, ikinci grupta konakçı iskelet sisteminde değişiklik yaparak bakterinin alınmasını sağlayan proteinler (SipB ve SipC) bulunur. SPA-1 ürünü olan Inv/Spa proteini makrofaj apoptozundan sorumlu iken; SipA aktin bağlayıcı bir proteindir. SopE ise GTP-bağlayıcı proteinleri aktive etmektedir. SPA-1'de yerleşim gösteren hilA da, transkripsiyonu düzenleyen proteinleri kodlamaktadır. Yapılan moleküler çalışmalar SPA-1'in, Salmonella enfeksiyonlarının enterik ateş (sistemik) veya yangılı ishallerin görüldüğü intestinal faz için önemli olduğunu göstermektedir4,7,12.
Salmonella Patojenite Adası-2 (SPA-2)
SPA-2, 40 kb büyüklüğünde olup, 32 geni kodlamaktadır. Bu genlerin çoğu konak hücre içinde bakteri üremesi sırasında eksprese edilir. Örneğin bunlardan biri olan spiC gen ürünleri, Salmonella içeren fagozomların lizozomla birleşmesini engellemektedir. Bu genin ekspresyonu sistemik enfeksiyon oluşumu için önemlidir ve bakterinin makrofaj içinde çoğalmasını kontrol eder4,7,13.
Salmonella Patojenite Adası-3 (SPA-3)
SPA-3, 17 kb büyüklüğündedir ve 10 gene sahiptir. Bu adacık S.enterica serovar Typhi ve Typhimurium ile S.bongori'de bulunur. SPA-3 tarafından kodlanan temel virülans faktörü, yüksek afiniteli magnezyum transport sistemi (MgtS)'dir. Sitoplazmik membranda yerleşim gösteren MgtS, Salmonella türlerinin gerek hücre içinde gerekse makrofajların içinde canlı kalabilmesi için gereklidir. MgtS, mgtC geni tarafından kodlanmakta ve Mg2+ naklini sağlamaktadır. Bu nedenle mutant serotipler MgtC eksikliği nedeniyle intraselüler olarak çoğalamazlar4,14.
Salmonella Patojenite Adası-4 (SPA-4)
SPA-4, 27 kb büyüklüğünde olup, tRNA'ya bitişik yerleşim göstermektedir. Bu adacık 18 genden oluşur ve Tip I sekresyon sistemi (TOSS)'ni kodlayan genleri içerdiği düşünülmektedir. SPA-4, sitotoksin sekresyonunda rol alır ve hücreyi apoptoza götürür. Bu genlerin kodladığı proteinler etkenin makrofaj içinde canlı kalmasını sağlar8,15,16.
Salmonella Patojenite Adası-5 (SPA-5)
Büyüklüğü 7.6 kb olan SPA-5 bölgesi 6 geni kodlamaktadır. SPA-5'in, SPA-1 ve SPA-2 tarafından kodlanan TTSS için gerekli efektör proteinlerin (SopB, SigD, PipB, vb) üretiminden sorumlu olduğu düşünülmektedir. SPA-5 daha ziyade Sop proteinlerini kodlayan genlerle karakterizedir. Örneğin TTSS'nin efektör bir proteini olan SopB bir inositol fosfataz olup, diyareye neden olan sıvı sekresyonunu tetikler. Bu nedenle SPA-5'in enterik enfeksiyonların patogenezinden sorumlu olduğu düşünülmektedir4,15,17.
Salmonella Patojenite Adası-6 (SPA-6)
SPA-6, 59 kb'lık bir bölgedir ve hem Typhi hem de Typhimurium serovarlarında bulunmaktadır. Bu adacık Salmonella kromozomal adacığı (SCI) olarak da bilinir ve fimbriyayı kodlayan saf geni, invazini kodlayan pagN geni ve fonksiyonu bilinmeyen birçok gen kümesini içerir. Yapılan çalışmalarda, serovar Typhimurium'daki SCI lokusu yok edildiğinde, etkenin neden olduğu sistemik enfeksiyonların patogenezinde bir değişiklik olmamakla birlikte, hücre kültüründe invazyonun gerçekleşmediği görülmüştür18,19.
Salmonella Patojenite Adası-7 (SPA-7)
Majör patojenite adası (MPI) olarak da bilinen SPA-7, 133 kb'lık en büyük Salmonella adacığıdır. SPA-7, serovar Typhi, Dublin ve Paratyphi C için özgül olup, diğer S.enterica serotiplerinde bulunmaz. Bu bölge aynı zamanda S.enterica serovar Typhi için MPI olarak da adlandırılır. Bu adacık tarafından kodlanan en önemli virülans faktörü Vi antijenidir. Vi antijeni kapsüler ekzopolisakkarit yapısında olup, tifo sırasında ateşin yükselmesine neden olmaktadır. Bir diğer virülans faktörü, pil gen kümesi tarafından kodlanan tip IVB pilusudur. SPA-7'de ayrıca sopE geni de bulunur. sopE geni sopE fajını aktive eder ve bu fajın diğer izolatlara transferini sağlar. sopE fajını da kodlayan SPI-7'nin genetik organizasyonu oldukça komplekstir. Bu lokus horizontal kazanılmış farklı elementlerden oluşmakta ve pil, tra ve sam genlerinin varlığı, SPA-7'nin bir konjugatif plazmid veya konjugatif transpozondan köken aldığını düşündürmektedir4,18,20,21.
Salmonella Patojenite Adası-8 (SPA-8)
SPA-8, 6.8 kb büyüklüğündedir ve pheV tRNA'nın yakınında yer alır. Bu adacık serovar Typhi için özgül olmakla birlikte, Saroj ve arkadaşları22, bu genin serovar Typhi dışında serovar Cholerasuis ve Paratyphi A'da da bulunduğunu bildirmişlerdir. SPA-8'de yer alan virülans faktörleri, bakteriyosin genleri tarafından sentezlenen bakteriyosinlerdir; ancak bu genlerin fonksiyonları henüz açıklık kazanmamıştır4,18,20.
Salmonella Patojenite Adası-9 (SPA-9)
SPA-9, yaklaşık 16 kb büyüklüğünde bir bölge olup, serovar Typhi kromozomundaki lizojenik bakteriyofaja bitişik olarak bulunur. SPA-9, TOSS ve RTX (repeats-in-toxin) benzeri toksini kodlayan genleri içermektedir4,18,20.
Salmonella Patojenite Adası-10 (SPI-10)
SPA-10, 32.8 kb büyüklüğündedir ve tRNA leuX'de yerleşim göstermektedir. SPA-10'da ayrıca kriptik bakteriyofaj da yer almaktadır. Bu lokus, serovar Typhi, Paratyphi A, Enteritidis, Dublin ve Gallinarum'da bulunmaktadır. SPA-10, virülans faktörü olarak, konak hücre yüzeyine tutunmayı sağlayan sef fimbriya proteinlerini kodlar18,20,22.
Salmonella Genomik Adası-1 (Salmonella Genomic Island; SGI-1)
SGI-1, çoklu direnç integronları içeren mobilize bir yapı olup, Salmonella enterica kökenlidir. Bir başka deyişle, çoklu antibiyotik direnç genlerini taşıyan gen kümelerini içerir. SGI-1, 43 kDa'luk bir lokus olup, kromozomal thdf ve int2 genleri arasında bulunur. Bu genomik ada Salmonella Typhimurium DT104, Paratyphi, Agona, Cerro, Derby, Dusseldorf, Emek, Haifa, Infantis, Kingston, Meleagridis, Newport, Typhimurium ve Virchow gibi pek çok S.enterica serotipinde bulunmaktadır. Bunlar arasında tıbbi önemi büyük olan S.Typhimurium DT104 suşu, dünyanın birçok bölgesindeki besin zehirlenmesi salgınlarından sorumludur. SGI-1 içeren suşların çoklu antibiyotik direnci (tetrasiklin, ampisilin, kloramfenikol, streptomisin ve sulfonamid direnci) gösterdiği saptanmıştır18,23,24.
Yüksek Patojenite Adası (High pathogenicity island; HPI)
Yüksek patojenite adası, demir alımı için ihtiyaç duyulan siderofor biyosentezi için gerekli genleri içerir ve S.enterica'da bulunur25. HPI'ya sahip olan suşlar demir eksikliği olan ortamlarda demir şelatörü sentezlemektedirler. Bu adacık ilk kez Yersinia türlerinde saptanmıştır ve birçok Enterobacteriaceae üyesinde yer almaktadır18.
TİP III SEKRESYON SİSTEMİ (TTSS)
"Moleküler enjektör" olarak da adlandırılan TTSS, temasa bağlı sekresyon veya konak hücreye virülans proteinlerin aktarımından sorumludur. TTSS, 20'den fazla proteinden oluşmuş kompleks bir yapı olup, bu proteinlerin organizasyonu enjektör (iğne kısmı, dış halka, boyun ve iç halka) yapısına benzemektedir. Bu sistem Salmonella dışında Shigella ve Escherichia coli'de de bulunur. TTSS'den sorumlu olan genler Salmonella'da kromozomal yerleşim göstermektedir. Salmonella enterica serotipleri SPA-1 ve SPA-2 tarafından kodlanan en az iki tip TTSS'ye sahiptirler. Bunlardan SPA-1 tarafından kodlanan sistem, intestinal enfeksiyonun başlatılmasından, SPA-2 tarafından kodlanan sistem ise sistemik enfeksiyonun oluşmasından sorumludur19,23,26.
VİRÜLANS GENLERİNİN REGÜLASYONU
Salmonella türlerinin başarılı bir enfeksiyona neden olabilmesi için, mide asiditesi, safra tuzu, oksijen kısıtlaması, besin yetersizliği, antimikrobiyal peptidlerin varlığı, mukus ve bağırsak içindeki doğal mikroflora gibi olumsuz çevre koşullarında canlı kalabilmesi gerekir. Salmonella'lar gerek invazyon gerekse makrofaj içinde çoğalmaları sırasında lizozomal enzimler, hidrojen peroksit, reaktif oksijen radikalleri, iNOS ve savunma sisteminin diğer elemanları gibi birçok olumsuz koşul ile karşılaşmaktadır. Bu nedenle bakterinin, enfeksiyon oluşturabilmesi için bir dizi gen ekspresyonuna gereksinimi vardır. İnvazyon ile ilişkili genler, 37°C'de, nötral pH'da, yüksek ozmolaritede ve üremenin geç döneminde maksimum düzeyde eksprese edilir ve SPA-1 tarafından kodlanan HilA (hyper-invasive locus) proteinleri tarafından regüle edilir. Virülan Salmonella suşları, fagozom veya fagolizozomlardaki olumsuz koşullara karşı varlıklarını "alternatif sigma faktörü" ve "adaptif aside tolerans yanıtı" ile sürdürmektedirler. Bu nedenle bu iki özellik bakterinin önemli virülans faktörlerindendir6,8,18,27.
Alternatif Sigma Faktör ss (RpoS) Regülatörü
RpoS, Salmonella türlerinin en önemli virülans faktörlerinden olup, besin yokluğu veya kısıtlaması ile pH ve sıcaklık değişimleri gibi stres faktörlerine maruz kaldığı zaman, bakterinin bu olumsuz koşullara dayanmasını ve canlı kalmasını sağlamaktadır28. Bu amaçla bakteri, durağan ve çoğalma fazında pek çok gen eksprese eder. Örneğin RpoS, Salmonella serovar Typhimurium'un derin organ bölgelerinde çoğalabilmesini kontrol eden spvRABCD plazmid genlerinin ekspresyonundan sorumludur. Bu genlerden spvB, makrofaj içinde bakterinin canlı kalmasını sağlayabilmek amacıyla ADP-ribozil-transferazı üretir29. Bu faktör ayrıca, Peyer plaklarında kolonizasyon için gerekli olan kromozomal genlerin fonksiyonlarını da düzenler30.
Adaptif Aside Tolerans Yanıtı
Normalde nötral pH'da yaşamaya alışkın olan Salmonella türleri, su birikintileri, mide asidi ve kolon gibi asidik çevre koşullarına da maruz kalabilirler. Aside tolerans yanıtı, özellikle bakterinin mideden gastrointestinal faza geçişi sırasında veya asidik fagozomal (pH: 5-6) çevre şartları içinde yaşamını sürdürebilmesi ve çoğalması için oldukça önemlidir. Fagozom, bakteriyi yok etmek için lizozom dışında, hidrojen peroksit, süperoksit ve diğer antimikrobiyal etkili proteinleri salgılarken, çeşitli metabolik aktiviteler sonucu fagolizozomun pH'sı 3-4'e kadar iner. Normalde bakteri, orta düzeyde asidik ortama (pH: 5) maruz kaldığında yanıt olarak bir dizi (~50 adet) gen ekspresyonu yapmaktadır. Bu genlerin ürünleri sayesinde bakteri daha asidik ortama (pH~3) adapte olur ve canlılığını sürdürür. RpoS, PhoP/Q, OmpR ve Fur gibi genel regülatör proteinler bu olaylar sırasında önemli rol oynarlar16,31,32. Aside toleranslı türlerin daha yüksek patojeniteye sahip olması nedeniyle aside tolerans yanıtı önemli virülans faktörlerinden biri olarak kabul edilmektedir.
KAYNAKLAR
- Tindall BJ, Grimont PA, Garrity GM, Euzeby JP. Nomenclature and taxonomy of the genus Salmonella. Int J Syst Evol Microbiol 2005; 55(Pt1): 521-4. [Özet] [Tam Metin] [PDF]
- Majowicz SE, Musto J, Scallan E, et al. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis 2010; 50(6): 882-9. [Özet] [Tam Metin] [PDF]
- World Health Organization. Global Salmonella Surveillence Database 2011. Erişim: http://www.who.int/gfn/general/documnts/GSS_STRATEGICPLAN2006_10.pdf
- Hensel M. Evolution of pathogenicity islands of Salmonella enterica. Int J Med Biol 2004; 291(2-3): 95-102. [Özet]
-
Schmidt H, Hensel M. Pathogenicity islands in bacterial pathogenesis. Clin Microbiol
Rev 2004; 17(1): 14-56.
[Özet] [Tam Metin] [PDF] - Ibarra JA, Steele-Mortimer O. Salmonella--the ultimate insider. Salmonella virulence factors that modulate intracellular survival. Cell Microbiol 2009; 11(11): 1579-86. [Özet] [Tam Metin] [PDF]
- Hall RM. Salmonella genomic islands and antibiotic resistance in Salmonella enterica. Future Microbiol 2010; 5(10): 1525-38. [Özet]
- Kaur J, Jain SK. Role of antigens and virulence factors of Salmonella enterica serovar Typhi in its pathogenesis. Microbiol Res 2012; 167(4): 199-210. [Özet]
- Bueno SM, Riquelme S, Riedel CA, Kalergis AM. Mechanisms used by virulent Salmonella to impair dendritic cell function and evade adaptive immunity. Immunology 2012; 137(1): 28-36. [Özet] [Tam Metin] [PDF]
-
Sabbagh SC, Forest CG, Lepage C, Leclerc JM, Daigle F. So similar, yet so different:
uncovering distinctive features in the genomes of Salmonella enterica
serovars Typhimurium and Typhi. FEMS Microbiol Lett 2010; 305(1): 1-13.
[Özet] [Tam Metin] [PDF] - Chiu CH, Tang P, Chu C, et al. The genome sequence of Salmonella enterica serovar Choleraesuis, a highly invasive and resistant zoonotic pathogen. Nucleic Acids Res 2005; 33(5): 1690-8. [Özet] [Tam Metin] [PDF]
- Layton AN, Galyov EE. Salmonella-induced enteritis: molecular pathogenesis and therapeutic implications. Expert Rev Mol Med 2007; 9(18): 1-17. [Özet]
- Figueira R, Holden DW. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology 2012; 158(Pt 5): 1147-61. [Özet] [Tam Metin] [PDF]
- Retamal P, Castillo-Ruiz M, Mora GC. Characterization of MgtC, a virulence factor of Salmonella enterica serovar Typhi. PLoS One 2009; 4(5): e5551. [Özet] [Tam Metin] [PDF]
- Sandra LM, Brumell JH, Pfeifer CG, Finlay BB. Salmonella pathogenicty islands: big virulence in small packages. Microbes Infect 2000; 2(2):145-56. [Özet]
- Altier C. Genetic and environmental control of salmonella invasion. J Microbiol 2005; 43(Spec No): 85-92. [Özet]
- Burkinshaw BJ, Prehna G, Worrall LJ, Strynadka NC. Structure of Salmonella effector protein SopB N-terminal domain in complex with host Rho GTPase Cdc42. J Biol Chem 2012; 287(16): 13348-55. [Özet]
- Morgan E. Salmonella pathogenicity islands, pp: 67-88. In: Rhen M, Maskell D, Mastroeni P, Threlfall J (eds), Salmonella: Molecular Biology and Pathogenesis. 2007, Horizon Bioscience. Norfolk, UK.
- Folkesson A, Lofdahl S, Normark S. The Salmonella enterica subspecies I specific centisome 7 genomic island encodes novel protein families present in bacteria living in close contact with eukaryotic cells. Res Microbiol 2002; 153(8): 537-45. [Özet]
- Parkhill J, Dougan G, James KD, et al. Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature 2001; 413(6858): 848-52. [Özet] [Tam Metin] [PDF]
- Seth-Smith HM. SPI-7: Salmonella's Vi-encoding pathogenicity island. J Infect Dev Ctries 2008; 2(4): 267-71. [Özet]
- Saroj SD, Shashidhar R, Karani M, Bandekar JR. Distribution of Salmonella pathogenicity island SPI-8 and SPI-10 among different serotypes of Salmonella. J Med Microbiol 2008; 57(Pt4): 424-7. [Özet] [Tam Metin] [PDF]
- Duard G, Praud K, Cloeckaert A, Doublet B. The Salmonella genomic island 1 is specifically mobilized in trans by the IncA/C multidrug resistance plasmid family. PLoS One 2010; 5(12): e15302. [Özet] [Tam Metin] [PDF]
- Mulvey MR, Boyd DA, Olson AB, Doublet B, Cloeckaert A. The genetics of Salmonella genomic island 1. Microbes Infect 2006; 8(7): 1915-22. [Özet]
-
Oelschlaeger TA, Zhang D, Schubert S, et al. The high-pathogenicity island is absent in
human pathogens of Salmonella enterica subspecies I but present in
isolates of subspecies III and VI. J Bacteriol 2003; 185(3): 1107-11.
[Özet] [Tam Metin] [PDF] - Schraidt O, Lefebre MD, Brunner MJ, et al. Topology and organization of the Salmonella Typhimurium type III secretion needle complex components. PLoS Pathog 2010; 6(4): e1000824. [Özet] [Tam Metin] [PDF]
- Alvarez-Ordóñez A, Begley M, Prieto M, et al. Salmonella spp. survival strategies within the host gastrointestinal tract. Microbiology 2011; 157(Pt 12): 3268-81. [Özet] [Tam Metin] [PDF]
-
Dong T, Schellhorn HE. Role of RpoS in virulence of pathogens. Infect Immun 2010;
78(3): 887-97.
[Özet] [Tam Metin] [PDF] -
Guiney DG, Fierer J. The role of the spv genes in Salmonella pathogenesis.
Front Microbiol 2011; 2:129.
[Özet] [Tam Metin] [PDF] - Nickerson CA, Curtiss R 3rd. Role of sigma factor RpoS in initial stages of Salmonella Typhimurium infection. Infect Immun 1997; 65(5): 1814-23. [Özet]
- Tiwari RP, Sachdeva N, Hoondal GS, Grewal JS. Adaptive acid tolerance response in Salmonella enterica serovar Typhimurium and Salmonella enterica serovar Typhi. J Basic Microbiol 2004; 44(2): 137-46. [Özet]
- Allam US, Krishna MG, Sen M, et al. Acidic pH induced STM1485 gene is essential for intracellular replication of Salmonella. Virulence 2012; 3(2): 122-35. [Özet] [Tam Metin] [PDF]
İletişim (Correspondence):
Doç. Dr. Belgin Sırıken,
Ondokuz Mayıs Üniversitesi Veteriner Fakültesi,
Besin Hijyeni ve Teknolojisi Anabilim Dalı,
Kurupelit Kampüsü, Samsun, Türkiye.
Tel (Phone): +90 362 312 1919-2813,
E-posta (E-mail): bsiriken@yahoo.com